INTRODUCTION
Polyneuropathy is a frequent complication of chronic consumption of ethanol, characterized by allodynia and pain, primarily in the lower extremities, and is poorly managed by available treatments22). It is often asymptomatic, and incidence of peripheral neuropathy ranges from 10% to 50%21). The mechanism of alcohol-induced neuropathy is an axonal neuropathy characterized by Wallerian degeneration of the axons and a reduction in the myelination of nerve fibers34).
The pathogenesis of this axonal degeneration and reduction in the myelination, however, is not well understood. Researches on animal models have suggested that alcohol has a direct toxic effect on peripheral nerve and the spinal cord system5,23). Especially acetaldehyde, one of the most important metabolites of ethanol, has a direct neurotoxic effect16). Epigenetic and inflammatory control is critical role in the experimental animal models of alcohol induced neuropathy. In the recent year, similar results have been obtained that protein kinase species play a critical role in the development and maintenance of alcohol-induced pain6). Extensive animal study suggested that chronic administration of minocycline could ameliorate the development of neuropathic pain by inhibiting the release of proinflammatory cytokines and oxidative stress in mononeuropathic rats7). A significant increase in lipid peroxidation and a significant decrease in the activity of antioxidant enzymes (superoxide dismutase and catalase) were observed in the sciatic nerves of diabetic rats with established neuropathic pain28). Dina et al.6) demonstrated that allodynia is present in an established rat model of chronic alcoholism and that an inflammatory process and protein kinase signaling play a pivotal role in the enhanced allodynia produced by chronic alcohol. An important action of cAMP is activation of transcription factors, including cAMP-responsive element binding (CREB) protein and nuclear factor-kB (NF-κB) p5012). Phosphorylation of CREB stimulates transcription of cell survival genes19). Phosphorylation of NF-κB p50 subunit suppresses transcription of inflammation-associated genes, especially proinflammatory cytokines tumor necrosis factor-α (TNF-α) and interleukin-1β (IL-Iβ)4,12,31). Thus, we hypothesized that use of rolipram, a selective inhibitor of cAMP specific phosphodiesterase (PDE), may improve mechanical allodynia and nerve conduction in a rat model of ethanol-induced neuropathic pain.
MATERIALS AND METHODS
Experimental animals
Male adult Sprague-Dawley rats weighing 200-300 g were used in this study. The animals were housed in groups of two in plastic cages with soft bedding and free access to food and water. All animals were acclimated to their cages for 1 week before any experiments were performed. All experimental protocols were approved by the Institutional Animal Care and Use Committee and were carried out in accordance with the National Institutes of Health's Guide for the Care and Use of Laboratory Animals.
Ethanol-induced neuropathic pain model and drug treatment protocol
Alcoholic neuropathy was induced by administering 10 g/kg b.i.d oral gavage of 35% v/v ethanol in double distilled water for 8 weeks in all animals. Rolipram (Sigma, St. Louis, MO, USA), 3 mg/kg was first dissolved in 75 µL and then gently mixed with 75 µL Tween 80 and 1850 µL physiological saline to a final 2,000 µL solution. Physiological saline served as the vehicle for the control group. According to the randomly assigned group, animals were administered rolipram or vehicle once daily for a period of eight weeks. 250 µL rolipram or vehicle was administered intraperitoneally. Intraperitoneal delivery involved an injection below the skin on the low abdominal region of the animal at the right flank.
Electrophysiologic responses
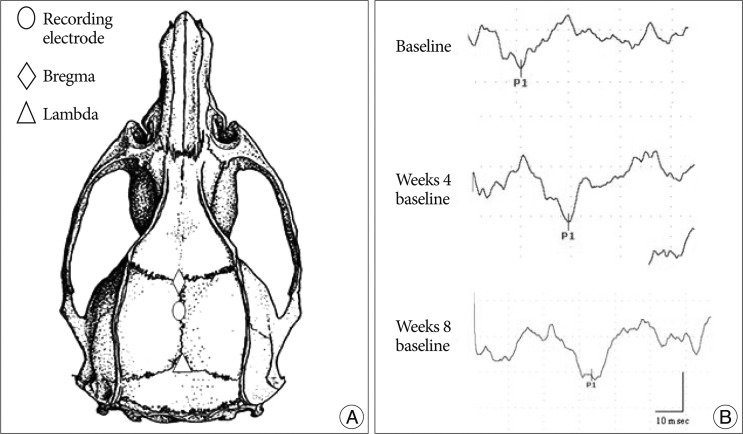
A Nicolet Viking IV was used for recording the somatosensory evoked potential (SEP). All SEP responses were obtained from stimulation of anesthetized, unrestrained rats. Rats were placed in a prone position on a plastic board with the active electrode implanted 2.5 mm posterior to the bregma and the reference electrode implanted in the mid frontal bone (Fig. 1). The SEP responses were elicited by activating the sensory nerve in the tale. For the sensory nerve stimulation, the cathode was placed 4 cm distal to the tale origin site and the anode was positioned two fingers distal to the cathode with ring electrode. The ground electrode was placed subcutaneously between the stimulation and recording electrodes. The tail was stimulated by positive 1.3-1.9 mA current pulses for 0.2 msec at 3 Hz. The signal-to-noise ratio was improved by ensemble averaging of 500 stimulus locked sweeps. The first peak latency was recorded. All animals were evaluated once a week for 8 weeks.
Quantification for mechanical allodynia
The behavioral tests measured were foot withdrawal thresholds (as an indicator for mechanical allodynia) in response to mechanical stimuli applied to the left and right hind paws22). The examiner who conducted the tests did not know about the nature of the experimental treatment. For each test, the animals were placed in a plastic chamber (9×9×30 cm) and habituated for at least 10 min. The chamber was placed on a mesh screen, so that mechanical stimuli could be administered to the plantar surface of the left and right hind paws. Thresholds were determined by the up-down method7) by using a set of von Frey monofilaments (von Frey filament values: 3.65, 3.87, 4.10, 4.31, 4.52, 4.74, 4.92, and 5.16; equivalent to: 0.45, 0.74, 1.26, 2.04, 3.31, 5.50, 8.32, and 14.45 g values). A von Frey filament was applied perpendicularly to the most sensitive areas of the plantar surface at the base of the third or fourth toes with sufficient force to bend the filament slightly for 3-4 sec. An abrupt withdrawal of the foot during stimulation or immediately after stimulus removal was considered as a positive response. The first stimulus was always the 4.31 filament. When there was a positive response, a filament with the next lower von Frey value was used. When no response was observed, a filament with the next higher von Frey value was applied. This testing pattern continued until responses to the sixth von Frey stimuli from the first change of response (either higher or lower than the first stimulus depending on whether the first response was negative or positive) were measured. The responses were then converted into a 50% threshold value using the formula: 50% threshold 10 (X
kd)/104, where X is the value of the final von Frey hair used in log units, k is the tabular value for the pattern of positive or negative responses, and d is the mean differences between stimuli in log units (0.22). When positive or negative responses were still observed with the 3.65 or 5.16 filament, values of 0.3 or 18.6 g were assigned, respectively, by assuming a value of 0.5 for k.
Statistical analysis
Results are presented as means±SEMs and analyzed using the SigmaStat program. Statistical analyses were done using two-way repeated-measures analysis of variances with two-repeated factors followed by Tukey post hoc test for the experiment of Latin square design or two-way repeated-measures analysis of variances with one repeated time factor followed by Tukey post hoc tests. In all cases, p<0.05 was considered significant.
RESULTS
SEP responses
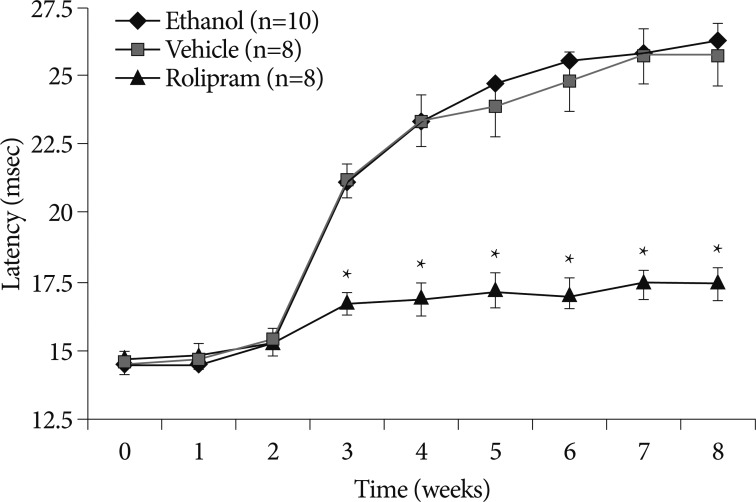
SEP study, a measure of sensory nerve excitability, was extensively used in the diagnosis of neuropathies. Beginning from week 3, latency significantly started to increase in control groups (rolipram 16.76±1.27, ethanol 21.10±1.75, vehicle 21.19±1.85) (p<0.05) (Fig. 3). In rolipram-treated rats, the shorter latency was sustained until week 8, the final week of the study (rolipram 17.50±1.77, ethanol 26.30±2.55, vehicle 25.80±3.17) (p<0.05) (Fig. 3).
Mechanical threshold
The baseline mechanical thresholds of all rats before ethanol application was 17.9 g which was the maximal cutoff point. Ethanol decreased the mechanical threshold of both hind paws. Further, the mechanical threshold of both hind paws did not significantly differ from each other. Thus, we used the left hind paw as the site to measure mechanical threshold. When ethanol (10 g/kg) was administrated on day 0 to week 8, there was a significantly greater decrease in nociceptive threshold, beginning from week 3 (rolipram 18.56±1.943, ethanol 4.370±1.631, vehicle 4.375±1.663) (p<0.05) (Fig. 2). The statistical difference was sustained until week 8, the final week of the study (rolipram 16.60±1.53, ethanol 1.23±0.45, vehicle 1.38±0.55) (p<0.05) (Fig. 2).
DISCUSSION
The neurologic effects of ethanol consumption are complex, affecting on both the central and peripheral nervous system26,32,35). In the peripheral nervous system, it produces a small-fiber dying back painful neuropathy2,6). Over time, pain far outweighs analgesia, producing a neuropathic syndrome with symptoms that have been described as "like tearing flesh off the bones"3). In recent years, ethanol consumption in Western industrialized countries has increased with an associated rise in the rates of alcohol-related health problems8,11). Thus, understanding the basis of clinical expression of ethanol-induced neuropathy is an issue of growing importance, and adequate treatment of this symptom may require therapeutic strategies.
Rolipram is a selective inhibitor of cAMP specific phosphordiesterase. PDE4 inhibitors have been reported to control epigenetic regulation and reduce both proinflammatory cytokine levels, including TNF-α and IL-1β1,25), molecules involved in free radical production and oxidative stress such as iNOS and COX-2, as well as immune cell infiltration into the nervous system28). Free radicals are derivatives of molecular oxygen and nitrogen and consist of superoxide, hydroxyl radical, hydrogen peroxide, and peroxynitrite17). These molecules are ubiquitously present in the body and participate in many normal cellular processes, including ion transport, transcription, neurotransmission, and neuromodulation17). Sources of free radicals are both mitochondrial oxidative metabolism which produces adenosine triphosphate and several enzymes such as xanthine oxidase, phospholipase A2, cytochrome P450, monoamine oxidase, and tyrosine hydroxylase. They are normally removed by antioxidant systems including superoxide dismutase, catalase, glutathione, glutathione peroxidase, ascorbate, and α-tocopherol. Thus, free radical levels are precisely controlled by antioxidant systems. However, in pathologic conditions, levels of free radicals may rise due to the increased production or decreased antioxidants level9,10).
Once a lesion develops in the central nervous system, two proinflammatory cytokines, TNF-α and IL-1β, are released. Numerous studies have documented rapid increases in TNF-α and IL-1β levels after nervous system injury such as traumatic brain injury and spinal cord injury occurs15,33). IL-1β synergistically acts with TNF-α to induce nerve cell death. These proinflammatory cytokines stimulate inflammatory cells to release damaging reactive oxygen and nitrogen species, raise glutamate levels to excitotoxic levels, impair the ability of glia cells to buffer extracellular potassium, compromise the nervous system and attract inflammatory cells into the nervous system13,14,20,29,30). Once initiated, the inflammatory cascade becomes a toxic positive-feedback loop, further exacerbating nervous system pathology. Several studies have demonstrated that restoration of cAMP levels improve outcome in models of nervous system injury. Rolipram which was used to inhibit the degradation of cAMP promoted axon sparing and improved functional outcomes24). The effects of cAMP are short-lived because they are rapidly degraded by PDEs18). Among ten PDE classes, two isoforms are highly selective for degrading cAMPs in the nervous system.
Our present study determined for the first time that ethanol-induced allodynia was significantly restored in the rolipram-treated rats compared with control groups consumed ethanol and received vehicle injection (Fig. 2, 3). Despite the impressive results of rolipram found in this study, cautious interpretation of study findings is warranted because of some limitation, such as study design without measurement of long-term effects and optimal dose of rolipram. Further study is needed to identify pathophysiology using cytokine analysis, ROS measurement and nerve biopsy.
CONCLUSION
This study shows that rolipram, a PDE4 inhibitor known to control epigenetic regulation and reduce both proinflammatory cytokine levels and molecules involved in free radical production and oxidative stress, ameliorated mechanical behavior as measured by mechanical allodynia and SEP in ethanol-induced neuropathy in rats.